
Download


Bladder | 2023 | Vol. 10 | e21200008
DOI: 10.14440/bladder.2023.870
DOI: 10.14440/bladder.2023.870
REVIEW
The Expression and Function of Piezo Channels in Bladder
Zhipeng Li1,2, Dongxu Lin1,2, Changcheng Luo1,2, Pengyu Wei1,2, Bolang Deng1,2, Kang Li1,2, Langqing Cheng1,2, Zhong Chen1,2*
1Department of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, Hubei, China
2Institute of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, Hubei, China
2Institute of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, Hubei, China
*Corresponding author: Zhong Chen, Department and Institute of Urology, Tongji Hospital, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, Hubei, China. Email: 1991tj0590@hust.edu.cn
Conflict of interest: No conflict of interest was declared by the authors.
Abbreviation used: [Ca2+]i, Intracellular calcium concentration; ATP, Adenosine triphosphate; BOO, Bladder outlet obstruction; CYP-induced, Cyclophosphamide-induced; DCP, Diabetic cystopathy; DRG, Dorsal root ganglia; FISH, Fluorescence in situ hybridization; GsMTx4, Grammostola spatulate mechanotoxin 4; ICCs, Interstitial Cajal-like cells; ICs, Interstitial cells; KO, Knockout; LP, Lamina propria ; LP-ICs, Interstitial cells in bladder lamina propria; LUT, Lower urinary tract; LUTS, Lower urinary tract symptoms; NCX1, Na+/Ca2+ exchanger 1; OAB, Overactive bladder; pBOO, Partial bladder outlet obstruction; PIEZO1-tdT, PIEZO1 fused to the red fluorescent reporter tandem-dimer Tomato; SAC, Stretch-activated ion channel ; SACs, Stretch-activated ion channels; TRPV4, Transient receptor potential vanilloid 4; UCs, Urothelial cells
Received July 19, 2023; Revision received September 15, 2023; Accepted October 9, 2023; Published October 30, 2023
Abstract
The ability for bladder to perceive and analyze mechanical stimuli, such as stretch and filling, is crucial for its functions, such as urinary storage and voiding. The Piezo channel family, including Piezo1 and Piezo2, represents one of the most essential mechanosensitive ion channels in mammals and is involved in a wide array of physiological and pathological processes. It has been demonstrated in numerous investigations that Piezo channels play a key role in mechanical transduction in various types of cells in bladder by converting mechanical stimuli into biological signals. Notably, mounting evidence suggests that Piezo channels are functionally significant for bladder and are related to several bladder disorders. This review systematically summarizes the importance/role and features of Piezo channels in bladder, including their biophysical properties, location, and functions, with attention specifically paid to their association with the physiology and pathophysiology of bladder. This review aims to provide a novel perspective for the future clinical treatment of bladder dysfunction.
Keywords: Bladder dysfunction, Mechanosensitive cation channels, Piezo channels
1 INTRODUCTION
The bladder is able to sense the mechanical stimuli generated by pressure and filling, and converts them into biological signals (including chemical signals and electrical signals), which are transmitted to the higher-level micturition center in the brain, allowing the brain to control urine storage and voiding in a coordinated manner[1,2]. Nonetheless, until several years ago, how mechanical stimuli are translated into electrical impulses in nerve fibers were poorly understood.
Mechanosensitive cation channels are an important class of mechanotransduction molecules and closely related to the transmission of mechanical signals in organisms[3,4]. They can adapt to the changes of mechanical stress on cell membrane by quickly changing their conformation, thereby converting the mechanical stimuli applied to the cell membrane into bioelectrical or biochemical signals[5]. Over a decade ago, Ardem Patapoutian and colleagues discovered a type of current signal activated by mechanical stimuli in a Neuro2A mouse neuroblastoma cell line. Afterwards, by knocking down candidate genes in cells respectively and measuring their responses to mechanical stimuli subsequently, researchers took over 3 years to identify the Piezo1 (Fam38A) and Piezo2 (Fam38B), two channels necessary for the mechanical activation of currents in those cells [5]. Further researches have shown that Piezo1 and Piezo2 are structurally comparable, both being capable of converting pressure applied to the surface of cell membrane into bioelectrical signals [5].
A multitude of physiological activities depend on the sensation and transmission of mechanical stimuli, such as touch and pain, the development of blood vessels, and the regulation of blood pressure[6,7]. In mammals, Piezo channels play a predominant role in mechanotransduction[5]. Since the discovery of the novel ion channels Piezo1/Piezo2 is crucial for the understanding of the mechanisms of mechanotransduction in mammals, Ardem Patapoutian and David Julies, who discovered the receptor for temperature, were jointly awarded the Nobel prices 2021 for Physiology and Medicine[8].
In this review, we summarize the studies regarding Piezo1/2 channels, including their biophysical properties, location and functions in bladder, and their association with the diseases of lower urinary tract. Through elucidating the role of Piezo1/2 channels in bladder, we tried to offer a novel perspective to approach the treatment and intervention of bladder disorders.
2 The biophysical properties and physiological roles of Piezo1/Piezo2
Mechanosensitive ion channel proteins are the molecular basis on which cells sense mechanical forces, and can convert mechanical stimuli into bioelectrical or biochemical signals in several milliseconds by changing their conformations [5]. This protein-mediated mechanotransduction represents one of the fastest conduction processes within living organisms [5]. Nevertheless, Piezo1 and Piezo2 are different with regard to functions, distribution patterns, and the way to be activated[4].
Piezo1, as one of the mechanosensitive ion channels, is activated by mechanical forces and converts them into electrical signals [9]. This Piezo1-mediated transduction from mechanical stimulus to electrical signal is very rapid [10]. Under physiological conditions, Piezo1 channel generates an inward current flowing into the cell, which leads to cellular depolarization and an increase of positive charges inside the cell. In theory, all types of physiological force that can change the tension on cell membrane are able to activate Piezo1 channel[11]. Besides, Piezo1 can respond to different forms of mechanical stimuli, such as puncturing, stretching, and fluid shear forces, among others[5]. Piezo1 does not require the participation of other cellular components to respond to mechanical forces, suggesting that the mechanosensitivity of Piezo1 lies in its intrinsic property and does not need signals from other proteins or second messengers[12]. The sensitivity of Piezo1 channel to lateral membrane tension is excellent. The resting membrane tension regulates the mechanical sensitivity of Piezo1 by inactivating the Piezo1 channel[13]. Piezo1 channel is one of the most representative non-selective cation channels, whose permeability to cations is in the following order: Ca2+> K+> Na+>Mg2+, with the permeability to Ca2+ being the highest[5]. The conductive ability of Piezo1 is related to its selective permeability to cations. In addition, studies have shown that extracellular matrix proteins increase the sensitivity of Piezo1 to mechanical forces. With the absence of extracellular matrix proteins, Piezo1 is relatively insensitive to mechanical forces exerted on the cell membrane, resulting in Piezo1 being almost unable to be activated by the mechanical stretching. The matrix gel, a mixture of extracellular matrix proteins, allows for the formation of a mechanical junction network between cells, which enhances the sensitivity of Piezo1 to mechanical stretching, thus enabling Piezo1 to be activated by mechanical forces more effectively[14]. In mammals, Piezo1 is mainly present in non-sensory tissues that are subject to fluid pressure or flow, such as renal, vesical, and cardiovascular tissues[15]. Moreover, Piezo1 plays an important role in the sensing of mechanical stimuli by cells, and is thereby implicated in a variety of pathological and physiological processes, such as vascular development, arterial remodeling, and blood pressure regulation[16].
Piezo2 is also a mechanosensitive ion channel that converts mechanical stimuli into electrical signals, and is involved in various pathological and physiological processes, such as gentle touch sensation, proprioception, mechanical hypersensitivity, and tumor angiogenesis[17]. Piezo2 is sensitive principally to low-threshold mechanical stimuli, mediating pain in mice and human and playing an important role in proprioception, cutaneous sensation, and visceral sensation in mammals. The permeability of Piezo2 channel to cations is in the following order: Ca2+> K+> Na+>Mg2+, which is similar to Piezo1[5,18]. In mammals, Piezo2 is mainly expressed in sensory tissues such as sensory neurons in dorsal root ganglia and Merkel cells, which mostly participates in the sensing of gentle touch, proprioceptive status, airway dilation, lung ventilation[11,15,19]. Deficiency in the expression of Piezo2 channel in sensory neurons and Merkel cells in mice leads to the loss of the ability to perceive gentle touch[20]. Similarly, after specific knocking Piezo2 out in neurons of dorsal root ganglia, mice lose sensitivity to gentle touch but still exhibit the response to noxious mechanical stimuli[21]. In the evaluation of inflammation or post-injury tissue state, the expression level of Piezo2 channel is often used as an indicator of the sensitivity of the body's response to mild mechanical stimuli[22]. In order to find the direct evidence that Piezo2 is a tactile receptor, in 2014, the Patapoutian laboratory conducted an investigation[20]. After applying mechanical stimuli to Merkel cells, mechanically-dependent currents were detected, which could lead to downstream action potentials, thus directly proving that Piezo2 is a tactile receptor. Another study in 2014 showed that knockout of Piezo2 gene in mice could cause severe loss of tactile sensation but did not impair other senses[22], which, from the behavioral perspective, also confirmed that Piezo2 is a tactile receptor . Additionally, Patapoutian laboratory observed that mice with the Piezo2 gene knocked-out exhibited severe lack of coordination in movements and abnormal sensation of limb position[23], thus substantiating that Piezo2 is the main receptor of proprioception in mice.
3 The location of Piezo1/Piezo2 in bladder
3.1 The location of Piezo1 in bladder
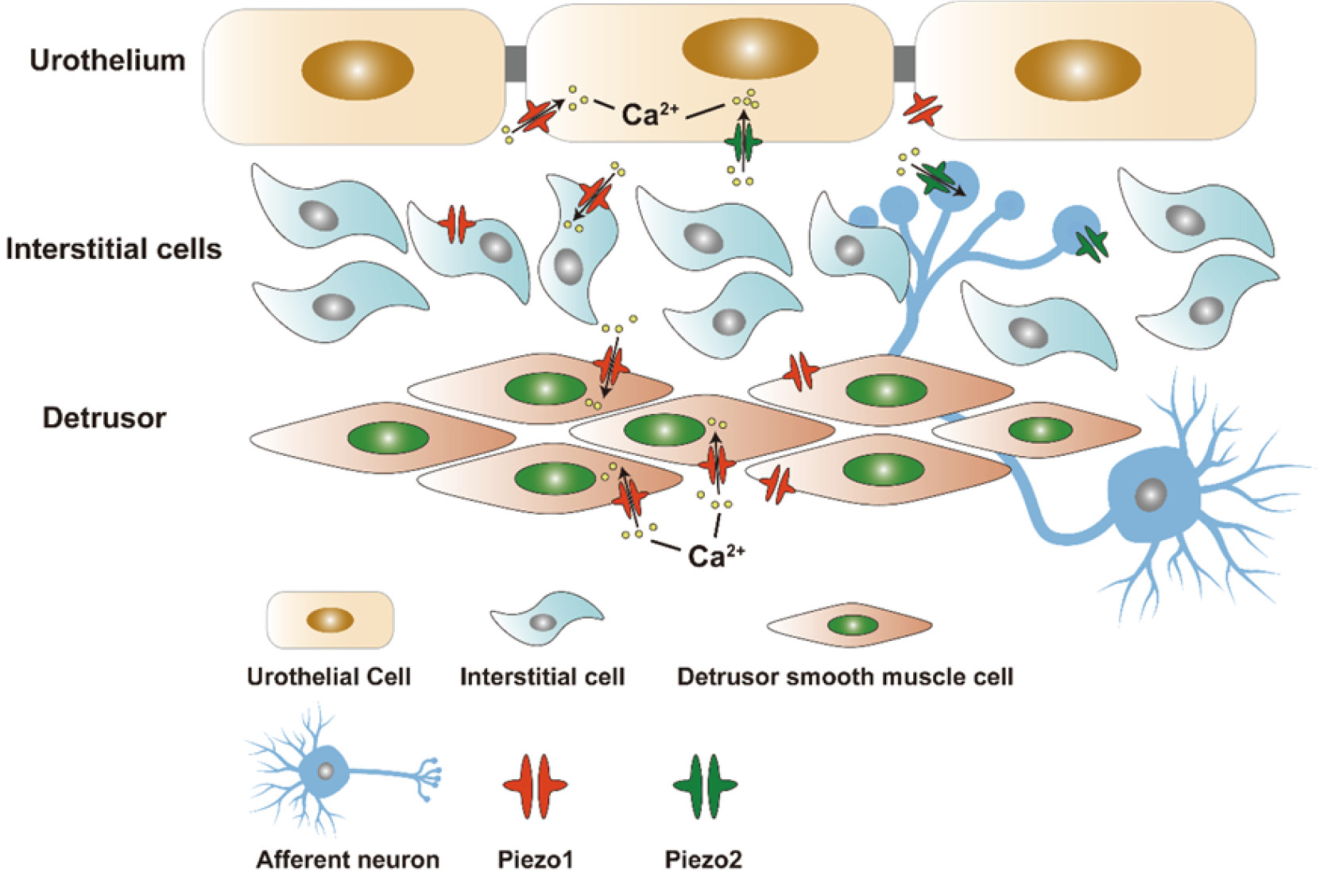
By examining transgenic mice which expressed PIEZO1 fused with the red fluorescent reporter tandem-dimer Tomato (PIEZO1-tdT), Dalghi et al. found that Piezo1 was expressed in kidney, ureter, bladder, urethra, and organs closely related to urinary tract, such as prostate gland, seminal vesicle, ejaculatory duct, and vagina. They also observed that Piezo1 was expressed at or in close proximity to the surfaces of the underlying intermediate and basal cell layers. Besides, Piezo1 was also found to express in the interstitial cells, smooth muscle cells and striated muscle cells of bladder and ureters[24]. The aforementioned findings suggest that Piezo1-tdT is present in the ureters and bladder, in which it is localized in the urothelial, mesothelial, and interstitial cells of the lamina propria, and smooth muscle cells forming the walls of these organs.
Dalghi and colleagues evaluated the expression of Piezo1 in murine urothelial cells by using fluorescence in situ hybridization (FISH). They confirmed that Piezo1 was expressed in all three layers of urothelia as well as in adjacent cells such as fibroblasts[17]. According to Dalghi, Piezo1 was localized at the basal and lateral surfaces of umbrella cells, and it was also found on the plasma membrane of intermediate cells and basal cells located beneath the umbrella cells (Fig.1). In native umbrella cells, Piezo1-tdT is distributed along the lateral membrane, but does not extend to tight junctions marked by CLDN8[24,25]. What is more, it was noted that Piezo1 is also expressed within cells situated deep in the walls of the ureter, bladder, and urethra[4].
Figure 1 The location of Piezo1/Piezo2 in bladder. Piezo1 is localized in the urothelium, interstitial cells of the lamina propria and detrusor smooth muscle cells. Piezo2 is observed in umbrella cells and afferent neurons.
3.2 The location of Piezo2 in bladder
Marshall et al. were the first to demonstrate that Piezo2 channel is expressed in more than 70% of umbrella cells, which form the innermost layer of the urothelium, and accounted for more than 80% of afferent neurons[26]. Besides, by using techniques such as fluorescence in situ hybridization (FISH), Dalghi et al. found that the mRNA of Piezo2 is mainly expressed in scattered clusters of umbrella cells in the bladder wall[17]. In addition, the expression of Piezo2 was also detected in suburothelial fibroblasts and cells in dorsal root ganglia [17,26].
4 The function of Piezo1/Piezo2 in bladder
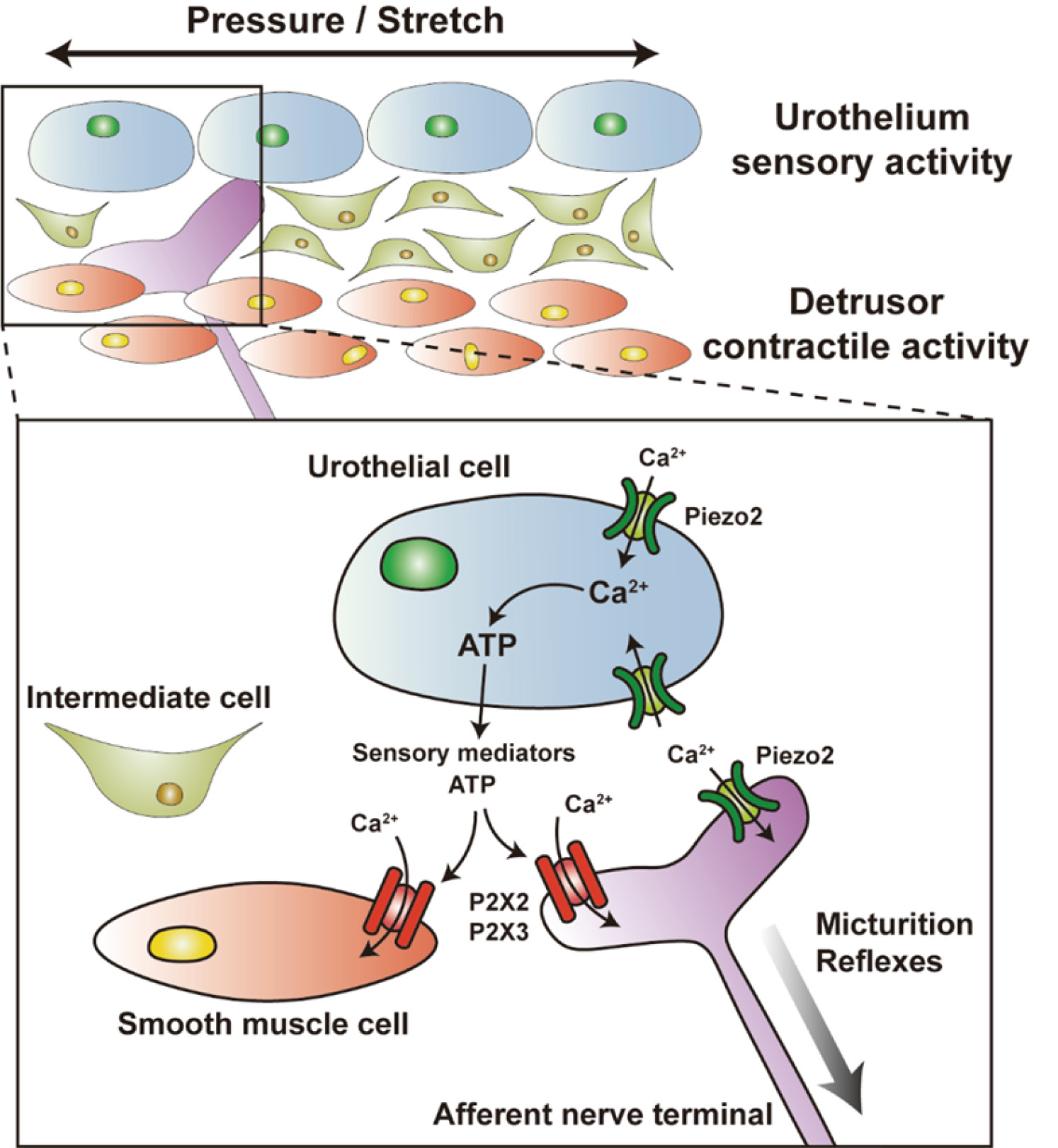
The urothelium of bladder, including umbrella cells, is believed to be a passive barrier against water, ions, solute molecules, and microbes that may cause infection[25]. However, the urothelium serves as not only a physical barrier, but also a detector that senses signals from bladder, including filling and voiding. It can sense the mechanical stretching of bladder wall, which leads to Ca2+ influx and release of adenosine triphosphate (ATP) in urothelial cells.[27,28]. ATP functions as a stimulant for afferent nerves in bladder wall, which interacts with P2X2, P2X3, and other purinergic receptors located on sensory nerve terminals. Subsequently, the signals about bladder filling level are transmitted to nerve center, where they trigger the micturition reflex and promote the voiding process.[29–31]. In a currently putative model, bladder filling leads to ATP release from urothelia, which is hypothesized to promote the afferent reflex in a stimulatory manner, or downstream micturition function of local urothelia (Fig.2)[32]. Prior studies have shown that suburothelial interstitial cells (ICs) in humans, when activated, express Piezo1 and release ATP. The interstitial cells in bladder lamina propria (LP-ICs) can sense/perceive mechanical stretching or chemical stimuli in bladder lamina propria (LP) via the activation or inactivation of TRPA1, TRPV2, TRPV4, and Piezo1. Besides, there may exist a bidirectional communication between LP-ICs and the surrounding urothelial cells or sensory afferent neurons through paracrine signaling, by which mechanical or chemical signals could be perceived and transduced, and thus the urothelium could play a role in the sensory function of bladder[33,34]. Furthermore, Piezo1 channel has also been detected in interstitial Cajal-like cells (ICCs) and detrusor smooth muscle cells, but, its biological roles in the urinary system have yet to be fully eluciated[16]. Piezo2 channel on umbrella cells and afferent nerves is thought to be critical for triggering the voiding reflex[23].
Figure 2 A hypothetical model describing the potential role of Piezo channels in the bladder wall. In response to mechanical stimuli induced by bladder wall distension, the mechanosensitive cation channels of urothelial cells, including Piezo1/2 and TRPV4, are activated. As a result of this process, Ca2+ enters the cells, triggering the exocytotic release of ATP. Afterwards, ATP binds to purinergic receptors (e.g., P2X2) located on the afferent nerve fibers, thereby conveying signals regarding urinary control to the central nervous system. The function of Piezo1 channel in interstitial cells and smooth muscle cells is still unclear. Piezo2 channel on afferent nerve fibers may be an important component for micturition reflex.
4.1 The function of Piezo1 in bladder
Piezo1, a subtype of stretch-activated ion channels (SACs)[35], is a mechanosensitive molecule expressed in urothelial cells and other types of cells of bladder in mice and humans[3,36]. It can be activated by the stretching of cell membrane, resulting in an increase in intracellular calcium concentration ([Ca2+]i), which then leads to ATP release. Utilizing siRNA targeting Piezo1 channel can suppress the cascade reactions induced by Ca2+ influx. Applying GsMTx4 to inhibit the function of Piezo1 results in a similar reduction in Ca2+ influx and the level of ATP release[36]. Miyamoto et al. reported that the release of ATP triggered by stretching in isolated mouse urothelial cells was reliant or partially reliant on Piezo1[36]. Subsequent studies indicated that ATP release in endothelial cells also relied on Piezo1[37]. In mice with conditional urothelial Piezo1 knockout, ATP release from the urothelium did not significantly decrease, and these mice exhibited normal voiding function and behaviors[17], which suggested that there might be a potential compensatory mechanism to overcome the deficiency of Piezo1. Nevertheless, the existing evidence does not suffice to prove whether overexpression of Piezo2 can act as a compensatory mechanism in response to the loss of Piezo1[17]. Transient receptor potential vanilloid 4 (TRPV4), as a stretch-activated ion channel (SAC), is expressed in the detrusor layer of bladder. Activation of TRPV4 could lead to Ca2+ influx, which could trigger bladder contraction subsequently[38,39]. Piezo1 may possess similar functions with TRPV4. In fact, the activation of Piezo1 in smooth muscle cells of bladder could cause Ca2+ influx[3,36], which can elicit bladder contraction. Furthermore, increased Piezo1 expression in the detrusor layer is able to enhance the bladder contractility activated by distension[40].
4.2 The function of Piezo2 in bladder
Marshall et al. demonstrated that Piezo2 functions as a mechanosensor in both urothelial cells and sensory neurons innervating the bladder (Fig.3). Besides, Piezo2 is necessary for sensing low-threshold bladder distension, which appears to participate in the relaxation of the detrusor muscle during bladder filling and the micturition reflex[26]. Nevertheless, though the essential roles of urothelia and the sensory neurons of bladder in mechanosensation have been established, further researches are required to comprehend the mechanisms by which sensitivity to pressure and distribution patterns of Piezo1/2, and signaling pathways activated by these channels contribute to the coordination of the voiding reflex.
Figure 3 The localization and physiological function of Piezo2 channel in bladder. The expression of Piezo2 channel (marked in green) can be observed in both umbrella cells and afferent nerve terminals (Aδ and C fibers). When exposed to pressure and stretch within bladder, umbrella cells are capable of releasing sensory mediators, including ATP, which can activate P2X2 and/or P2X3 receptors (marked in red). Both urothelial cells and afferent neurons are able to sense stretch and/or pressure, ultimately activating the micturition reflex.
Marshall and colleagues monitored the bladder pressure and sphincter activity in UPKII-Cre+ Piezo2fl/fl mice, which had been subjected to an extensive knockout of Piezo2 in their urothelial cells. Mice with Piezo2 knocked-out demonstrated higher threshold for sensation to bladder stretching, requiring higher bladder pressure to initiate micturition, and exhibited a weakened urethral reflex[26]. By utilizing different mouse models with Piezo2 conditionally knocked-out, Marshall et al. discovered that, in these mice, the neurons located in S1 dorsal root ganglia exhibited, in general, a decreased sensitivity to bladder distension, and, in particular, an absence of sensitivity to pressures less than 20 mmHg. Cystometry coupled with urethral electromyography revealed that mice with Piezo2 knockout had reduced sensitivity to bladder distension and experienced irregular micturition timing, longer intervals between bladder voiding contractions, higher bladder pressure just prior to those micturition contractions, and larger voiding volumes[18,26]. Premised on these findings, it can be inferred that the reflex relaxation of the detrusor muscle in response to bladder filling was disturbed, or bladder compliance might be compromised in the absence of Piezo2. As the mice with Piezo2 knockout aged, they developed detrusor hypertrophy, an indication of chronic bladder dysfunction. Moreover, impaired coordinated activity of the urethra suggested that Piezo2 is responsible for the sensitivity of the lower urinary tract to stretching and triggers proper micturition reflexes[18]. In order to distinguish between the functions of Piezo2 in umbrella cells and sensory neurons, Marshall et al. knocked out Piezo2 in Aδ and C fibers specifically[26]. Mice with neuron-specific Piezo2 knockout showed longer intervals between bladder contractions, but no significant difference in pre-contraction pressure compared with wild-type mice, which further highlighted the primary role of Piezo2 in initiating bladder relaxation in umbrella cells. However, mice with neuron-specific Piezo2 knockout required higher bladder pressure for voiding and showed weakened urethral contraction, indicating a critical role of Piezo2 in/on neurons in these reflexes[26,41]. Despite reports of voiding abnormalities in both mice with Piezo2 knockout and patients with Piezo2 defects/deficiency, their urinary function and behaviors are not completely impaired.[26]. It is important to note that Piezo2 also appears to contribute to normal micturition in humans. Chesler reported 12 patients with Piezo2 defects/deficiency who mostly, apart from sensory dysfunction, exhibited reduced frequency of urination and episodes of sudden urge incontinence[42].
4.3 The synergistic function of Piezo1 and Piezo2 in bladder
During bladder filling, ATP is released from urothelial cells, which conveys the level of bladder distension to subjacent tissues, including afferent nerve fibers, and activates the voiding reflex subsequently (Fig.2). In order to investigate whether Piezo channels play a role, as mechanosensors, in these events, Dalghi et al. produced murine models with conditional knockout (KO) of Piezo1, Piezo2, and both Piezo1/2 specifically in urothelial cells respectively[17]. Although Piezo1 channel is present across all cell layers of the urothelium, the phenotype of Piezo1-knockout mice was limited. Urethane-anesthetized Piezo1-control and Piezo1-knockout mice underwent continuous cystometrical testing. In female mice with loss of Piezo1, no significant effect of Piezol absence on cystometric parameters was observed. Similarly, no significant difference was observed in cystometrical parameters between male Piezo1-KO mice and controls. Owing to technical difficulties in collecting urine from male mice, their voiding efficiency could not be measured accurately. The expression of Piezo2 is confined to a minor population of superficial umbrella cells. Mice with conditional urothelial Piezo2-KO exhibited normal mechanotransduction and lower urinary tract (LUT) function, but male mice with Piezo2 channel knocked-out showed urinary incontinence (i.e., leakage) during their active dark phase, while no significant effect on voiding efficiency was observed in female Piezo2-KO mice. Mice with dual Piezo1/2 knockout exhibited the most pronounced phenotype, characterized by reduced response to mechanical stimuli and diminished ATP release. It was observed that anesthetized female dual Piezo1/2-KO mice exhibited bladder hypoactivity, but their male counterparts did not. Besides, during the active dark phase, but not the inactive light phase, male and female mice with dual Piezo1/2-KO suffered from urinary incontinence. Dalghi proposed that urothelia function in a sex-dependent and circadian fashion, and established a connection between mechanotransduction mediated by urothelial Piezo1/2 channels and normal micturition function and behaviors. In these studies, knockout of Piezo1 or Piezo2 in urothelial cells alone was found to be insufficient to effect changes, whereas mice with both Piezo1 and Piezo2 channels knocked-out presented symptoms of urinary incontinence [17], which indicated that simultaneous knockout of both Piezo1 and Piezo2 channels is necessary to reveal the true phenotypes.
5 Piezo channels are involved in diseases of lower urinary tract
As a mechanotransduction mediator, abnormalities of Piezo channels are associated with various clinical diseases, including cystitis, bladder outlet obstruction, overactive bladder, nocturia, diabetic cystopathy, and bladder cancer.
5.1 Cystitis
Liu et al. investigated the functional role of Piezo1 channel in cyclophosphamide-induced (CYP-induced) cystitis. After treatment with CYP for 48 h or 8 d (CYP-48h or CYP-8d), the expression of Piezo1 channel was significantly increased in interstitial Cajal-like cells (ICCs) of bladder, with a more significant increase in CYP-8d rats. Bladder hyperactivity in CYP-8d rats was significantly alleviated by treatment with the peptide toxin grammostola spatulate mechanotoxin 4 (GsMTx4), an antagonist of Piezo1 channel. Additionally, by using GsMTx4 and siRNA targeting Piezo1 channel, researchers demonstrated that Piezo1 channel was significantly activated during chronic cystitis induced by CYP, which evoked Ca2+ and Na+ influx in ICCs of bladder. Furthermore, in ICCs of CYP-8d rats, the functional interaction was observed between Piezo1 channel and the relatively activated reverse mode of Na+/Ca2+ exchanger 1 (NCX1) [16].
On the basis of relevant researches, Liu et al. proposed that Piezo1 channel was easily activated during bladder filling or by cell swelling in the condition of cystitis. The activation of Piezo1 channel promotes Ca2+ influx, leading to Ca2+ overload in bladder ICCs, which may be a key pathogenic factor underlying bladder hyperactivity associated with chronic cystitis. Simultaneously, in chronic cystitis induced by CYP, Piezo1 channel synergizes with the reverse mode of NCX1 to greatly elevate the concentration of intracellular Ca2+ ([Ca2+]i) in ICCs, which then leads to Ca2+ overload (Fig. 4). This suggests that Piezo1 channel and NCX1 are potential new therapeutic targets of bladder hyperactivity associated with chronic cystitis[16].
Figure 4 Piezo1 channel interacts with the reverse mode of NCX1 in ICCs. In chronic cystitis induced by cyclophosphamide, Piezo1 channel in interstitial Cajal-like cells (ICCs) of bladder can be activated by mechanical stimuli, which leads to the influx of Ca2+ and Na+. Simultaneously, through the reverse mode of Na+/Ca2+ exchanger 1 (NCX1), Na+ from cytoplasm is extracted in exchange for additional Ca2+ entering ICCs. The synergistic effects of the Piezo1 channel and NCX1 increased the intracellular concentration of Ca2+, which results in Ca2+ overload in bladder ICCs. This may be a potential mechanism underlying bladder hyperactivity related to chronic cystitis.
Moreover, studies by Liu et al. confirmed that the expression of Piezo1 was increased significantly in the suburothelial layer during chronic cystitis[16]. In the detrusor region, the expression of Piezo1 was markedly elevated in interstitial cells situated in the intermuscular region, but showed no significant alteration in detrusor muscle cells [16].
5.2 Bladder outlet obstruction (BOO)
Bladder outlet obstruction (BOO) is characterized by elevated resistance in the urinary outflow tract. BOO results from different diseases, such as benign prostatic hyperplasia in males, bladder neck obstruction in females and urethral stricture[43,44]. Clinically, lower urinary tract symptoms are the primary signs of BOO, which involve storage symptoms (such as nocturia, urinary frequency, and urgency) and voiding symptoms (such as feelings of incomplete voiding, weak stream, hesitancy)[45]. However, the exact pathological mechanisms underlying bladder dysfunction associated with the obstructive diseases mentioned above have not been completely elucidated. It has been reported that bladder hyperactivity in mice with partial bladder outlet obstruction (pBOO) is related to the downregulated expression of mechanosensitive TREK-1 potassium channel in the detrusor muscle[46]. Another investigation suggested that detrusor overactivity in rats with BOO was associated with TRPV4[47].
Michishita et al. created a rat model of pBOO by partial urethral ligation[40], and observed a significant increase in Piezo1 mRNA in the suburothelial and detrusor layers of the rat bladder on the 7th day after pBOO. Besides, with the mRNA of Piezo1 increasing, the expression of neurofilaments dropped[40]. Piezo1 expressed in suburothelial layer may participate in the transduction of signals to afferent nerve endings, mimicking its function in the urothelium. Consequently, the upregulated Piezo1 expression in the suburothelial layer in rats with pBOO could potentially increase the sensation of bladder to distension and micturition. In contrast, the mechanism by which Piezo1 in detrusor muscle layer contributes to bladder dysfunction may differ from that at work in the suburothelial and urothelial layers. Interestingly, the activation of TRPV4, which is expressed in the detrusor muscle and urothelium, causes bladder contractions that are not dependent on the urothelium [38]. Therefore, Piezo1 in the detrusor muscle may directly induce bladder contractions, and the increased expression of Piezo1 in the detrusor muscle may also enhance the contractility of the bladder. In summary, the increased expression of Piezo1 in the bladder with pBOO may be implicated in bladder dysfunction through neurogenic and myogenic mechanisms.
5.3 Overactive bladder (OAB)
In recent years, the urothelial-derived mechanism has been incrementally recognized, which has opened up new perspectives for the study of overactive bladder (OAB). This theory suggests that the urothelium, as a non-neuronal internal sensor, may lead to overactive bladder (OAB) when inappropriately stimulated by bladder pressure and chemical substances[48–52]. Growing evidence indicates that changes in the expression and sensitivity of the mechanoreceptor in urothelia may affect the sensitivity of bladder to mechanical stretching, which could result in abnormal bladder activity [31,53].
As researches on Piezo channels continue to move forward, more reliable evidence has been found to support the urothelial-derived mechanism. Mechanical stimuli, such as stretching of the bladder wall, result in the elevation of intracellular Ca2+ concentration ([Ca2+]i) in the urothelium. This process is mediated by Piezo1/2, and subsequently lead to ATP release.[17,36]. It has been reported virtually all ATP released from serosa depends on mechanotransduction mediated by Piezo1 or Piezo2[17]. Additionally, researchers have observed a marked elevation of Piezo1 expression in both the urothelium and interstitial cells of Cajal (ICCs) of rats with cystitis[16]. Urodynamic measurement of and contractility testing on isolated bladder strips from rats demonstrated that GsMTx4 was able to alleviate bladder overactivity effectively[16]. Furthermore, patients with Piezo2 defects showed bladder underactivity[17]. However, mice with dual urothelial Piezo1/2 knockout developed hyperactive bladder, even urinary incontinence, instead of hypoactive bladder [17]. Therefore, the current evidence seems to contradict the straightforward conclusion that the upregulation of Piezo1/2 channels expression in the urothelium could result in overactive bladder (OAB). It is possible that changes in biophysical properties of Piezo channels, such as alterations in inactivation properties, may occur under pathological conditions.
Obviously, further researches are warranted to assess whether the mechanotransduction mediated by Piezo1/2 is altered under the condition of bladder hyperactivity. Remarkably, a previous study demonstrated that targeted knockdown of Piezo2 in bladder sensory neurons of murine models could result in prolonged intervals between bladder contractions and higher intravesical pressure needed to trigger micturition[26], suggesting that downregulating the expression of Piezo2 in bladder sensory neurons may hold therapeutic potential for managing overactive bladder (OAB).
5.4 Nocturia
Voiding function of bladder is under the control of clock genes with a circadian rhythm. The primary function of the bladder is micturition during the day, ensuring regular and efficient voiding frequency, while at night the chief function is storage of urine, which promotes better rest and sleep[54–56]. The disruption of the circadian rhythmic homeostasis of bladder function can lead to diseases such as nocturia.
Recent reports have suggested that the time-dependent pattern of gene expression is applicable to Piezo1. Therefore, the function of the urothelium perceiving bladder filling may possess a circadian rhythm[57]. Specifically, in mice, the expression level of Piezo1 in the urothelium rises during the active phase and declines during the sleep phase[58]. In addition, it was observed that the mutation of the clock gene is able to maintain a constant expression level of Piezo1 in the bladder[58]. The oscillation of Ca2+ influx mediated by Piezo1 in urothelial cells of mice also exhibits a circadian rhythm, which is consistent with the expression level of the Piezo1 channel and might be a crucial reason why the bladder perception of filling has a circadian rhythm[57]. Chromatin immunoprecipitation experiments have yielded evidence that the clock protein is able to bind to the promoter of Piezo1, and regulate its gene transcription subsequently [58]. More importantly, there exists a relationship between the levels of the expression of the circadian clock gene and Piezo1, and the observed damping effect of GsMTx4 on urine sensation in wild-type mice. And the effect is greatest during the sleeping phase[59]. On the strength of the findings, modifying the circadian rhythm of voiding function by inhibiting Piezo1 channel may potentially become a novel therapeutic strategy for nocturia. However, it's worth noting that the inhibitory effect of GsMTx4 on Piezo1 channel is non-specific, which also affects other mechanosensitive ion channels[59]. Therefore, this factor must be taken into consideration in jumping to the conclusion that GsMTx4 is beneficial to the treamtment of Piezo1-related diseases.
5.5 Diabetic cystopathy (DCP)
After being activated, Piezo1 can promote hyperplasia of bladder mucosa, leading to bladder hypertrophy, which is considered to be one of the mechanisms by which diabetic cystopathy (DCP) results in bladder hypertrophy [60]. Zhang et al. found that, at the early stage of DCP, the expression of Piezo1 and Piezo2 in bladder significantly increased, rendering the bladder more sensitive to mechanical stimulation, which results in lower urinary tract symptoms (LUTS), such as urinary frequency. At the later stage of DCP, the expression of Piezo1 and Piezo2 in bladder shows no significant difference as compared with the control group. Subsequently, it was observed that the bladder sensitivity to mechanical stimulation decreased, which eased the symptom of urinary frequency but increased the volume of residual urine. This phenomenon may be ascribed to the damage of bladder tissue caused by hyperglycemia. So far, no study examined the signaling pathway of Piezo channels in DCP and further research is needed[61].
5.6 Bladder cancer
Across the globe, bladder cancer is the second most common malignancy of the urinary system, with most cases being urothelial carcinomas[62]. Etem and colleagues investigated the expression levels of Piezo1/2 in bladder carcinoma in both humans and mice[63]. They collected tissue samples from 60 bladder carcinoma patients, while bladder carcinoma in mice was induced by N-butyl-N-(4-hydroxybutyl) nitrosamine (BBN). By using reverse transcription PCR and immunohistochemistry, they compared the expression levels of Piezo1/2 between normal tissues and bladder cancer tissues from human subjects and mice respectively. PCR showed that the expression of Piezo1/2 in bladder cancer tissues (both human and mice) was significantly upregulated in comparison with normal tissues. Besides, the expression of piezo1 in bladder cancer was found to bear a significant correlation with the size and the stage of tumor, but not with other clinicopathological features. On the other hand, the expression of Piezo2 was only associated with the stage of tumor, with higher expression level of Piezo2 observed in high-grade bladder carcinoma. In addition, no significant difference was found in the expression of Piezo1/2 between bladder cancer samples with lymph node metastasis and those without[63]. However, further investigations are warranted as the functions of Piezo1/2 channels in bladder cancer have not been fully elucidated [63,64].
6 Conclusion
As a mechanoreceptor, Piezo1/2 channels are closely associated with the sensing and transmission of mechanical signals in mammals. In recent years, there has been a significant increase in the number of studies on Piezo channels[65]. In the urinary system, Piezo channels have been proved to play a key role in mechanical transduction in different types of tissues. In addition, Piezo channels are widely distributed in various types of cells in the bladder, which can convert the mechanical stimuli applied to bladder into bioelectrical or biochemical signals, thus partaking in a variety of physiological and pathological processes. Furthermore, as a mechanotransduction mediator, the abnormality of Piezo channels is related to various clinical bladder disorders, such as overactive bladder and nocturia. Therefore, in the area of clinical application, Piezo channels might potentially emerge as a novel target for bladder dysfunction treatment, particularly for patients afflicted with complicated bladder dysfunction. In comparison to conventional therapeutic approaches, selective drugs and specific strategies targeting Piezo channels may be efficacious for patients with overactive bladder (OAB) or other bladder dysfunctions, for whom the efficacy of muscarinic cholinoceptor blockers (e.g., tolterodine and solifenacin) or β-adrenoceptor agonists (e.g., mirabegron) proves suboptimal or unsatisfactory[64]. In summary, although there is still a long way to go to reveal the exact mechanisms by which Piezo channels regulate the bladder function, Piezo channels, undoubtedly, are a very promising therapeutic target for bladder diseases.
Funding source
This work was supported by a grant from the National Natural Science Foundation of China (grant number 82270812).
Clinical trial registration and ethical approval:
References
- Vanneste M, Segal A, Voets T, Everaerts W. Transient receptor potential channels in sensory mechanisms of the lower urinary tract. Nat Rev Urol. 2021 Mar;18(3): -139–159. doi:10.1038/s41585-021-00428-6 https://doi.org/10.1038/s41585-021-00428-6 PMID: 33536636
- Griffiths D. Neural control of micturition in humans: a working model. Nat Rev Urol. 2015(Dec);12:695–705.doi:10.1038/nrurol.2015.266 https://doi.org/10.1038/nrurol.2015.266 PMID: 26620610
- Bagriantsev SN, Gracheva EO, Gallagher PG. Piezo proteins: regulators of mechanosensation and other cellular processes. J Biol Chem. 2014 Nov;289(46):31673–81. https://doi.org/10.1074/jbc.R114.612697 PMID: 25305018
- Li X, Hu J, Zhao X, Li J, Chen Y. Piezo channels in the urinary system. Exp Mol Med. 2022 Jun;54(6): 697–710. doi:10.1038/s12276-022-00777-1 https://doi.org/10.1038/s12276-022-00777-1 PMID: 35701561
- Coste B, Mathur J, Schmidt M, Earley TJ, Ranade S, Petrus MJ, et al. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. 2010Oct;330(6000): 55–60. https://doi.org/10.1126/science.1193270 PMID: 20813920
- Savadipour A, Palmer D, Ely EV, Collins KH, Garcia-Castorena JM, Harissa Z, et al. The role of PIEZO ion channels in the musculoskeletal system. Am J Physiol Cell Physiol. 2023 Mar;324(3):C: 728–40. https://doi.org/10.1152/ajpcell.00544.2022 PMID: 36717101
- Zeng W-Z, Marshall KL, Min S, Daou I, Chapleau MW, Abboud FM, et al. PIEZOs mediate neuronal sensing of blood pressure and the baroreceptor reflex. Science. 2018 Oct;362(6413):464–467.10.1126/science.aau6324 https://doi.org/10.1126/science.aau6324 PMID: 30361375
- Reeh PW, Fischer MJM. Nobel somatosensations and pain. Pfluger Arch. 2022Apr;474(4): 405–20. https://doi.org/10.1007/s00424-022-02667-x PMID: 35157132
- Lin YC, Guo YR, Miyagi A, Levring J, MacKinnon R, Scheuring S. Force-induced conformational changes in PIEZO1. Nature. 2019;573: 230–34. https://doi.org/10.1038/s41586-019-1499-2 PMID: 31435018
- Servin-Vences MR, Richardson J, Lewin GR, Poole K. Mechanoelectrical transduction in chondrocytes. Clin Exp Pharmacol Physiol. 2018May;45(5): 481–88. https://doi.org/10.1111/1440-1681.12917 PMID: 29359488
- Murthy SE, Dubin AE, Patapoutian A. Piezos thrive under pressure: mechanically activated ion channels in health and disease. Nat Rev Mol Cell Biol. 2017Dec;18(12): 771–83. https://doi.org/10.1038/nrm.2017.92 PMID: 28974772
- Syeda R, Florendo MN, Cox CD, Kefauver JM, Santos JS, Martinac B, et al. Piezo1 Channels Are Inherently Mechanosensitive. Cell Rep. 2016Nov;17(7): 1739–46. https://doi.org/10.1016/j.celrep.2016.10.033 PMID: 27829145
- Lewis AH, Grandl J. Mechanical sensitivity of Piezo1 ion channels can be tuned by cellular membrane tension. eLife. 2015Dec;4: e12088. https://doi.org/10.7554/eLife.12088 PMID: 26646186
- Gaub BM, Müller DJ. Mechanical Stimulation of Piezo1 Receptors Depends on Extracellular Matrix Proteins and Directionality of Force. Nano Lett. 2017Mar;17(3): 2064–2072. https://doi.org/10.1021/acs.nanolett.7b00177 PMID: 28164706
- Wu J, Lewis AH, Grandl J. Touch, Tension, and Transduction – The Function and Regulation of Piezo Ion Channels. Trends Biochem Sci. 2017Jan;42(1): 57–71. https://doi.org/10.1016/j.tibs.2016.09.004 PMID: 27743844
- Liu Q, Sun B, Zhao J, Wang Q, An F, Hu X, et al. Increased Piezo1 channel activity in interstitial Cajal-like cells induces bladder hyperactivity by functionally interacting with NCX1 in rats with cyclophosphamide-induced cystitis. Exp Mol Med. 2018May;50(5): 1–16. https://doi.org/10.1038/s12276-018-0088-z PMID: 29735991
- Dalghi MG, Ruiz WG, Clayton DR, Montalbetti N, Daugherty SL, Beckel JM, et al. Functional roles for PIEZO1 and PIEZO2 in urothelial mechanotransduction and lower urinary tract interoception. JCI Insight. 2021Oct;6(19): e152984. https://doi.org/10.1172/jci.insight.152984 PMID: 34464353
- Gailly P, Devuyst O. PIEZO2, a mechanosensor in the urinary bladder. Kidney Int. 2021Jul;100(1): 9–11. https://doi.org/10.1016/j.kint.2021.02.021 PMID: 33667505
- Wang Y, Chi S, Guo H, Li G, Wang L, Zhao Q, et al. A lever-like transduction pathway for long-distance chemical- and mechano-gating of the mechanosensitive Piezo1 channel. Nat Commun. 2018Apr;9(1): 1300. https://doi.org/10.1038/s41467-018-03570-9 PMID: 29610524
- Woo SH, Ranade S, Weyer AD, Dubin AE, Baba Y, Qiu Z, et al. Piezo2 is required for Merkel-cell mechanotransduction. Nature. 2014May;509(7502): 622–26. https://doi.org/10.1038/nature13251 PMID: 24717433
- Kim SE, Coste B, Chadha A, Cook B, Patapoutian A. The role of Drosophila Piezo in mechanical nociception. Nature. 2012Feb;483(7388): 209–12. https://doi.org/10.1038/nature10801 PMID: 22343891
- Ranade SS, Woo SH, Dubin AE, Moshourab RA, Wetzel C, Petrus M, et al. Piezo2 is the major transducer of mechanical forces for touch sensation in mice. Nature. 2014Dec;516(7529): 121–25. https://doi.org/10.1038/nature13980 PMID: 25471886
- Woo SH, Lukacs V, de Nooij JC, Zaytseva D, Criddle CR, Francisco A, et al. Piezo2 is the principal mechanotransduction channel for proprioception. Nat Neurosci. 2015Dec;18(12): 1756–1762. https://doi.org/10.1038/nn.4162 PMID: 26551544
- Dalghi MG, Clayton DR, Ruiz WG, Al-bataineh MM, Satlin LM, Kleyman TR, et al. Expression and distribution of PIEZO1 in the mouse urinary tract. Am J Physiol-Renal Physiol.2019Aug;317(2):F303–21. https://doi.org/10.1152/ajprenal.00214.2019 PMID: 31166705
- Dalghi MG, Montalbetti N, Carattino MD, Apodaca G. The Urothelium: Life in a Liquid Environment. Physiol Rev. 2020Oct;100(4): 1621–705. https://doi.org/10.1152/physrev.00041.2019 PMID: 32191559
- Marshall KL, Saade D, Ghitani N, Coombs AM, Szczot M, Keller J, et al. PIEZO2 in sensory neurons and urothelial cells coordinates urination. Nature. 2020Dec;588(7837): 290–95. https://doi.org/10.1038/s41586-020-2830-7 PMID: 33057202
- Dunning-Davies BM, Fry CH, Mansour D, Ferguson DR. The regulation of ATP release from the urothelium by adenosine and transepithelial potential: Adenosine and ATP release from the urothelium. BJU Int. 2013Mar;111(3): 505–13. https://doi.org/10.1111/j.1464-410X.2012.11421.x PMID: 22882496
- Mochizuki T, Sokabe T, Araki I, Fujishita K, Shibasaki K, Uchida K, et al. The TRPV4 cation channel mediates stretch-evoked Ca2+ influx and ATP release in primary urothelial cell cultures. J Biol Chem. 2009 Aug;284(32):21257–64. https://doi.org/10.1074/jbc.M109.020206 PMID: 19531473
- Aresta Branco MS, Gutierrez Cruz A, Peri LE, Mutafova-Yambolieva VN. The Pannexin 1 Channel and the P2X7 Receptor Are in Complex Interplay to Regulate the Release of Soluble Ectonucleotidases in the Murine Bladder Lamina Propria. Int J Mol Sci. 2023 Jun;24(12):9964. https://doi.org/10.3390/ijms24129964 PMID: 37373111
- Wei L, Mousawi F, Li D, Roger S, Li J, Yang X, et al. Adenosine Triphosphate Release and P2 Receptor Signaling in Piezo1 Channel-Dependent Mechanoregulation. Front Pharmacol. 2019Nov;10: 1304. https://doi.org/10.3389/fphar.2019.01304 PMID: 31780935
- Vlaskovska M, Kasakov L, Rong W, Bodin P, Bardini M, Cockayne DA, et al. P2X3 knock-out mice reveal a major sensory role for urothelially released ATP. J Neurosci. 2001 Aug;21(15):5670–7. https://doi.org/10.1523/JNEUROSCI.21-15-05670.2001 PMID: 11466438
- Birder L, Andersson KE. Urothelial Signaling. Physiol Rev. 2013Apr;93(2):653-80. https://doi.org/10.1152/physrev.00030.2012 PMID: 23589830
- Kefauver JM, Ward AB, Patapoutian A. Discoveries in structure and physiology of mechanically activated ion channels. Nature. 2020Nov;587(7835): 567–76. https://doi.org/10.1038/s41586-020-2933-1 PMID: 33239794
- Zhao M, Chen Z, Liu L, Ding N, Wen J, Liu J, et al. Functional Expression of Transient Receptor Potential and Piezo1 Channels in Cultured Interstitial Cells of Human-Bladder Lamina Propria. Front Physiol. 2022Jan;12: 762847. https://doi.org/10.3389/fphys.2021.762847 PMID: 35069237
- McCubbin S, Jeoung A, Waterbury C, Cooper RL. Pharmacological profiling of stretch activated channels in proprioceptive neurons. Comp Biochem Physiol Part C Toxicol Pharmacol. 2020Jul;233:108765. https://doi.org/10.1016/j.cbpc.2020.108765 PMID: 32305458
- Miyamoto T, Mochizuki T, Nakagomi H, Kira S, Watanabe M, Takayama Y, et al. Functional Role for Piezo1 in stretch-evoked Ca2+ Influx and ATP release in urothelial cell cultures. J Biol Chem. 2014Jun;289(23): 16565–16575. https://doi.org/10.1074/jbc.M113.528638 PMID: 24759099
- Wang S, Chennupati R, Kaur H, Iring A, Wettschureck N, Offermanns S. Endothelial cation channel PIEZO1 controls blood pressure by mediating flow-induced ATP release. J Clin Invest. 2016Dec;126(12): 4527–4536. https://doi.org/10.1172/JCI87343 PMID: 27797339
- Skryma R, Prevarskaya N, Gkika D, Shuba Y. From urgency to frequency: facts and controversies of TRPs in the lower urinary tract. Nat Rev Urol. 2011Oct;8(11): 617–30. https://doi.org/10.1038/nrurol.2011.142 PMID: 21971315
- Thorneloe KS, Sulpizio AC, Lin Z, Figueroa DJ, Clouse AK, McCafferty GP, et al. N-((1S)-1-{[4-((2S)-2-{[(2,4-dichlorophenyl)sulfonyl]amino}-3-hydroxypropanoyl)-1-piperazinyl]carbonyl}-3-methylbutyl)-1-benzothiophene-2-carboxamide (GSK1016790A), a novel and potent transient receptor potential vanilloid 4 channel agonist induces urinary bladder contraction and hyperactivity: Part I. J Pharmacol Exp Ther. 2008Aug;326(2): 432–442. https://doi.org/10.1124/jpet.108.139295 PMID: 18499743
- Michishita M, Yano KI, Tomita K, Matsuzaki O, Kasahara K. Piezo1 expression increases in rat bladder after partial bladder outlet obstruction. Life Sci. 2016Dec;166: 1–7. https://doi.org/10.1016/j.lfs.2016.10.017 PMID: 27756599
- Merrill L, Gonzalez EJ, Girard BM, Vizzard MA. Receptors, channels, and signalling in the urothelial sensory system in the bladder. Nat Rev Urol. 2016Apr;13(4): 193–204. https://doi.org/10.1038/nrurol.2016.13 PMID: 26926246
- Chesler AT, Szczot M, Bharucha-Goebel D, Čeko M, Donkervoort S, Laubacher C, et al. The Role of PIEZO2 in Human Mechanosensation. N Engl J Med. 2016Oct;375(14): 1355–1364. https://doi.org/10.1056/NEJMoa1602812 PMID: 27653382
- Wang W, Ai J, Liao B, Xiao K, Lin L, Chen H, et al. The roles of MCP-1/CCR2 mediated macrophage recruitment and polarization in bladder outlet obstruction (BOO) induced bladder remodeling. Int Immunopharmacol. 2021Oct;99: 107947. https://doi.org/10.1016/j.intimp.2021.107947 PMID: 34311189
- Metcalfe PD, Wang J, Jiao H, Huang Y, Hori K, Moore RB, et al. Bladder outlet obstruction: progression from inflammation to fibrosis. BJU Int. 2010Dec;106(11): 1686–1694. https://doi.org/10.1111/j.1464-410X.2010.09445.x PMID: 20590549
- Gratzke C, Bachmann A, Descazeaud A, Drake MJ, Madersbacher S, Mamoulakis C, et al. EAU Guidelines on the Assessment of Non-neurogenic Male Lower Urinary Tract Symptoms including Benign Prostatic Obstruction. Eur Urol. 2015Jun;67(6): 1099–1109. https://doi.org/10.1016/j.eururo.2014.12.038 PMID: 25613154
- Baker SA, Hatton WJ, Han J, Hennig GW, Britton FC, Koh SD. Role of TREK-1 potassium channel in bladder overactivity after partial bladder outlet obstruction in Mmouse. J Urol. 2010Fen;183(2): 793–800. https://doi.org/10.1016/j.juro.2009.09.079 PMID: 20022044
- Cho KJ, Park EY, Kim HS, Koh JS, Kim JC. Expression of transient receptor potential vanilloid 4 and effects of ruthenium red on detrusor overactivity associated with bladder outlet obstruction in rats. World J Urol. 2014jun;32(3): 677–682. https://doi.org/10.1007/s00345-013-1099-y PMID: 23700140
- Wiseman OJ, Fowler CJ, Landon DN. The role of the human bladder lamina propria myofibroblast. BJU Int. 2003Jan;91(1): 89–93. https://doi.org/10.1046/j.1464-410X.2003.03802.x PMID: 12614258
- Birder LA, de Groat WC. Mechanisms of Disease: involvement of the urothelium in bladder dysfunction. Nat Clin Pract Urol. 2007Jan;4(1): 46–54. https://doi.org/10.1038/ncpuro0672 PMID: 17211425
- Hawthorn MH, Chapple CR, Cock M, Chess-Williams R. Urothelium-derived inhibitory factor(s) in¯uences on detrusor muscle contractility in vitro. Br J Pharmacol. 129.
- Lee SR, Kim HJ, Kim A, Kim JH. Overactive bladder is not only overactive but also hypersensitive. Urology. 2010 May;75(5):1053–9. https://doi.org/10.1016/j.urology.2009.10.045 PMID: 20092879
- Peyronnet B, Mironska E, Chapple C, Cardozo L, Oelke M, Dmochowski R, et al. A Comprehensive Review of Overactive Bladder Pathophysiology: On the Way to Tailored Treatment. Eur Urol. 2019Jun;75(6): 988–1000. https://doi.org/10.1016/j.eururo.2019.02.038 PMID: 30922690
- Gevaert T, Vriens J, Segal A, Everaerts W, Roskams T, Talavera K, et al. Deletion of the transient receptor potential cation channel TRPV4 impairs murine bladder voiding. J Clin Invest. 2007Nov;117(11): 3453–62. https://doi.org/10.1172/JCI31766 PMID: 17948126
- Ramsay S, Zagorodnyuk V. Role of circadian rhythms and melatonin in bladder function in heath and diseases. Auton Neurosci. 2023 May;246:103083. https://doi.org/10.1016/j.autneu.2023.103083 PMID: 36871511
- Herrera GM, Meredith AL. Diurnal Variation in Urodynamics of Rat. Yamazaki S, editor. PLoS ONE. 2010;5: e12298. https://doi.org/10.1371/journal.pone.0012298.
- Parsons M, Tissot W, Cardozo L, Diokno A, Amundsen CL, Coats AC, et al. Bladder Diary Research Team Normative bladder diary measurements: Night versus day: Normative Day and Night Bladder Diary Measurements. Neurourol Urodyn. 2007;26(4): 465–473. https://doi.org/10.1002/nau.20355 PMID: 17335055
- Ihara T, Mitsui T, Nakamura Y, Kanda M, Tsuchiya S, Kira S, et al. The oscillation of intracellular Ca2+ influx associated with the circadian expression of Piezo1 and TRPV4 in the bladder urothelium. Sci Rep. 2018Apr;8(1): 5699. https://doi.org/10.1038/s41598-018-23115-w PMID: 29632308
- Ihara T, Mitsui T, Nakamura Y, Kira S, Nakagomi H, Sawada N, et al. Clock Genes Regulate the Circadian Expression of Piezo1, TRPV4, Connexin26, and VNUT in an Ex Vivo Mouse Bladder Mucosa. Bartell PA, editor. PLOS ONE. 2017;12: e0168234. https://doi.org/10.1371/journal.pone.0168234
- Ihara T, Mitsui T, Shimura H, Tsuchiya S, Kanda M, Kira S, et al. Different effects of GsMTx4 on nocturia associated with the circadian clock and Piezo1 expression in mice. Life Sci. 202Aug 1;278: 119555. https://doi.org/10.1016/j.lfs.2021.119555 PMID: 33930366
- Gudipaty SA, Lindblom J, Loftus PD, Redd MJ, Edes K, Davey CF, et al. Mechanical stretch triggers rapid epithelial cell division through Piezo1. Nature. 2017Mar;543(7643): 118–21. https://doi.org/10.1038/nature21407 PMID: 28199303
- Kun Z, Fang K, Fei Y, Bo Y, Li-Jun Y, Fu-Li W, et al. Effects of diabetes mellitus on the Piezo ion channels in rat bladder tissues. J Mod Urol. 2018.
- Compérat E, Amin MB, Cathomas R, Choudhury A, De Santis M, Kamat A, et al. Current best practice for bladder cancer: a narrative review of diagnostics and treatments. The Lancet. 2022Nov;400(10364): 1712–1721. https://doi.org/10.1016/S0140-6736(22)01188-6 PMID: 36174585
- Etem E, Ceylan GG, Özaydın S, Ceylan C, Özercan I, Kuloğlu T. The increased expression of Piezo1 and Piezo2 ion channels in human and mouse bladder carcinoma. Adv Clin Exp Med. 2018Aug;27(8): 1025–31. https://doi.org/10.17219/acem/71080 PMID: 30010255
- De Felice D, Alaimo A. Mechanosensitive Piezo Channels in Cancer: Focus on altered Calcium Signaling in Cancer Cells and in Tumor Progression. Cancers. 2020Jul;12(7): 1780. https://doi.org/10.3390/cancers12071780 PMID: 32635333
- Guo J, Gu D, Zhao T, Zhao Z, Xiong Y, Sun M, et al. Trends in Piezo Channel Research Over the Past Decade: A Bibliometric Analysis. Front Pharmacol. 2021Apr;12: 668714. https://doi.org/10.3389/fphar.2021.668714 PMID: 33935792